The West Indian Snipe was described in 2016, its fossil or subfossil remains were recovered from Late Pleistocene to Early Holocene deposits on several islands and island groups including the Bahamas, the Cayman Islands, Cuba, and the Turks and Caicos Islands.
The West Indian Snipe was a volant species but probably had quite short wings, this can be assumed from the shape of its wing bones. [1][2]
***
The species probably died out for more or less natural reasons, following changing environmental conditions due to changes in the height of the sea level after the beginning of the Holocene. The last remaining populations were then probably extirpated by the first Amerindian settlers.
********************
References:
[1] William Suárez: The enigmatic snipe Capella sp. (Aves: Scolopacidae) in the fossil record of cuba. Caribbean Journal of science 40(1): 155-157. 2004 [2] David W. Steadman; Oona M. Takano: A new extinct species of Snipe (Aves: Scolopacidae: Gallinago) from the West Indies. Zootaxa 4109(3): 345-358. 2016
The Bahamian Giant Owl, described in 1936 based on subfossil remains, was formerly thought to have been endemic to the Bahaman islands but is now known to have also occurred on the island of Cuba.
It was the largest species within its genus, and its remains are by far the rarest to be found, especially on Cuba.
The Bahamian Giant Barn Owl is known to have preferably preyed upon the Bahamian Hutia (Geocapromys ingrahami (J. A. Allen)), a large, nocturnal rodent species that still inhabits some of the Bahamian islands today. [2]
***
At least two additional species can be added to this list. Tyto pollens was a flightless, 1-m-tall congener of barn owls that likely occurred in old-growth Caribbean pine (Pinus caribaea) barrens of Andros Island, Bahamas, in association with early human settlers (see figs. 6-9). It probably gave rise to local lore of chickcharnies, a mischievous leprechaunlike, nocturnal imp said to have three toes and the ability to turn its head all the way around. If disturbed, chickcharnies would impart terrible misfortune. It is possible that territorial defense behaviors of a meter-tall Tyto could give rise to such legends. [1]
*********************
References:
[1] Bruce G. Marcot: Owls of old forests of the world. General Technicl Reports. Portland, Oregon: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station 1-64. 1995 [2] William Suárez; Storrs L. Olson: Systematics and distribution of the giant fossil barn owls of the West Indies (Aves: Strigiformes: Tytonidae). Zootaxa 4020 (3): 533-553. 2015
The Bahaman Caracara was described in 1959 based on fossil or subfossil bones, the oldest can be dated to a Pleistocene age, however, some are of Holocene origin and are only several thousand years old and even contain nearly complete mitochondrial DNA. [1]
The species inhabited the islands of the Bahamas as well as Cuba, it reached asize of about 60 cm, it had short wings and appears to have had only weak flying abilities.
The Bahaman Caracara clearly disappeared due to human interference.
*********************
References:
[1] Jessica A. Oswald; Julia M. Allen; Kelsey E. Witt; Ryan A. Folk; Nancy A. Albury; David W. Steadman; Robert P. Guralnick: Ancient DNA from a 2,500-year-old Caribbean fossil places an extinct bird (Caracara creightoni) in a phylogenetic context”. Molecular Phylogenetics and Evolution. 140: 106576. doi:10.1016/j.ympev.2019.106576. 2019
Originally, Brace’s Emerald was only ever known by a single male specimen that had been collected in 1877 on the island of New Providence, Bahamas; this was long ignored completely and was considered identical with the Cuban Emerald (Chlorostilbon ricordii (Gervais)) which also inhabits the Bahamas. In 1945 it was then considered to be a subspecies of the Cuban Emerald; only in 1987 it was recognized as having been a completely distinct species.
Brace’s Emerald is now also known from fossil bones that were recovered from Pleistocene deposits on New Providence in the 1980s; it is now understood as a Pleistocene relict that had survived into modern times only to disappear completely after its discovery.
*********************
References:
[1] Gary R. Graves; Storrs L. Olson: Chlorostilbon bracei Lawrence, an extinct species of hummingbird from new Providence Island, Bahamas. The Auk 104: 296-302. 1987
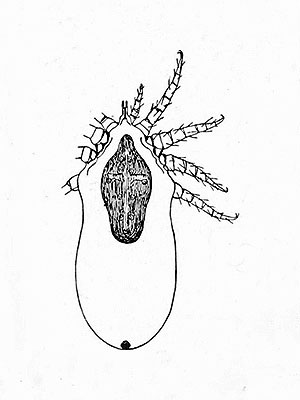
Caribbean Monk Seal Nasal Mite (Halarachne americana)
The Caribbean Monk Seal Nasal Mite was a parasite, specialized to the now extinct Caribbean Monk Seal (Neomonachus tropicalis (Gray)) where they lived inside the respiratory tract of that seal species, their biology, however, is not fully understood yet.
The mites disappeared together with their host species when the Caribbean Monk Seal died out in the 1950s.
***
This genus contains at least two additional species which are still alive, the Grey Seal Nasal Mite (Halarachne halichoeri Allman) and the Hawaiian Monk Seal Nasal Mite (Halarachne laysanae Furman & Dailey), both apparently restricted to their own single seal species host.
*********************
female
Depiction from: ‘Nathan Banks: A treatise on the Acarina, or mites. Proceedings of the United States National Museum 28: 1-114. 1905’
Crooked Island Hutia (Geocapromys ingrahami ssp. irrectus)
The Bahaman Hutia (Geocapromys ingrahami (J. A. Allen)) is a species of middle-sized rodent that is endemic to the Bahamas, or at least to the islands that are part of the so-called Great Bahama Bank.
The species was believed to be extinct, but a very small population was rediscovered in 1966 on a likewise very small island named East Plana Cay, this was assigned to the nominate form and in 1973, some of these animals were released on two additional islands, Little Wax Cay and Warderick Wells.
Based on DNA studies it is now known that the animals on East Plana Cay in fact did not constitute the last surviving wild population but one that was introduced in pre-Columbian times by the people of the Lucayan culture, the first human inhabitants of the Bahamas. [2]
***
Subfossil remains of this species were found on many of the larger islands, including Abaco as well as Crooked Island, Eleuthera, Exuma and Long Island; these forms differed from the alleged nominate form by their cranial characters and thus were described as two distinct subspecies: Geocapromys ingrahami ssp. abacoensis Lawrence (from Abaco Island) and Geocapromys ingrahami ssp. irrectus (from the remaining islands); the form from Abaco Island, however, is now known to have also just been introduced in prehistoric times, thus has never been a distinct subspecies. [1]
The animals that inhabited Crooked Island, Eleuthera, Exuma and Long Island on the other hand, appear to be non-monophyletic, thus likewise do not represent a distinct subspecies but either more than one or none at all.
The above-mentioned differences in the cranial characters may in fact just be the consequence of resource availability – animals on larger islands may have become larger because they had access to larger resources …. [2]
***
The Bahamian Hutia, once believed to be extinct, then rediscovered in 1966, may never had have any subspecies after all. This question is not yet answered – I will mention this species here only fro the sake of completness and because it constitutes a very interesting case.
*********************
References:
[1] B. N. Lawrence: Geocapromys from the Bahamas. Occasional papers of the Boston Society of Natural History 8: 189-196. 1934 [2] Jessica A. Oswald; Julie M. Allen; Michelle J. LeFebvre; Brian J. Stucky; Ryan A. Folk; Nancy A. Albury; Gary S. Morgan; Robert P. Guralnick; David W. Steadman: Ancient DNA and high-resolution chronometry reveal a long-term human role in the historical diversity and biogeography of the Bahamian hutia. Scientific Reports 10: 1373. 2020
*********************
Depiction from: ‘Mark Catesby; George Edwards: The natural history of Carolina, Florida, and the Bahama Islands : containing the figures of birds, beasts, fishes, serpents, insects, and plants : particularly, those not hitherto described, or incorrectly figured by former authors, with their descriptions in English and French : to which is prefixed, a new and correct map of the countries, with observations on their natural state, inhabitants, and productions. London: printed for B. White 1771’